





Жизнь глазами экономиста: эмпирические парадигмы развития и роста
Феномен жизни по-прежнему остается загадкой. С какой стати живые организмы усложняются? Зачем живым системам накапливать избыточные элементы в своей структуре? Есть ли глубинная аналогия между биомолекулярными и экономическими процессами? Что лежит в основе роста систем?
1. Предварительные замечания. С этой разработкой ознакомились известный биолог Александр Марков, физик Валерий Эткин и математик Дмитрий Абраров. Автор ведет в ней полемику с академиком Э.М.Галимовым и его известным произведением «Феномен жизни» [1]. Статья посвящена поиску внутренних размерностей биологического развития (роста), которые сближают этот процесс с пониманием динамики экономического и технологического прогресса. К этой экзотической проблеме как бы приложено мышление профессионального экономиста.
Предложена и анализируется эмпирическая модель поступательного «шагового» фактора биологического развития (step-процесс). Она исходит из представления о том, что биологическая эволюция имеет внутренний источник развития, действующий в форме тенденции к спонтанному росту биополимеров, начиная с ранних молекулярных форм преджизни. Эта тенденция проявляется на всем протяжении эволюции, определяя облик биосферы земли. Данная концепция относится к той группе воззрений, которые «мирно уживаются» с дарвиновским отбором, но вносят в него дополнительный смысл.
В то же время, по мнению А.В.Маркова, дарвиновское объяснение жизни является вполне достаточным и эффективным, и нет никакой необходимости в «лишних идеях» (бритва Оккама).
Статья носит характер интеллектуального эксперимента, своего рода проверки научной логики и эмпирического мышления. Она предполагает некоторое общее знакомство читателя с полемикой вокруг термодинамических, конкурентных и других начал эволюции.
2. Введение. Откажемся сразу от банальностей типа «эффективность», «оптимальное распределение ресурсов» или «рыночная конкуренция». Ниже пойдет речь о процессе, который ближе всего соответствует термину вроде «капитализации живого вещества», поскольку речь идет о природе ее структурной сложности. В то же время экономист стал бы, вероятно, искать фундаментальный фактор, своего рода избыточный ресурс, обеспечивающий прогрессивную эволюцию жизни. При этом он учел бы не только полезность «продукции» и текущую конъюнктуру, но и «ожидания рынка», его общие перспективы. Для этого рассмотрим жизнь как обобщенный объект – в пределах доступных нам описаний.
Метод исследования будет заключаться в поиске когнитивных измерителей, структурных шкал основных биологических процессов, обобщенной логики, на которую не могут «решиться» профильные специалисты. Мы предложим недоказанные допущения, вытекающие из общего обзора проблемы. Мы их утрируем и расширяем, поскольку для того, чтобы концепцию можно было проверить, ее следует попытаться в достаточной степени развить, что и составляет основу этого эксперимента. Здесь необходим контроль за экономичностью и очевидным смыслом введенных «новых сущностей», их взаимодействию и связью с классическим нормативом.
3. Биология vs. термодинамика. Наш сценарий опирается также на идеи термодинамики, но в ограниченном смысле, поскольку можно говорить лишь о некоторых аналогиях. Мы довольно рискованно введем «недостающее звено» – процесс, который еще не описали, и проверим, как может выглядеть картина в целом, если бы такой процесс действительно существовал.
Политико-экономические корни этого подхода заключаются в «эмпирическом шовинизме», энергичном конструировании реальности средствами эмпирических полутеорий, а также макроскопическом рассмотрении проблем подобно тому, как феномен промышленно-экономического роста обобщается в виде тенденции к развитию, вытекающей из свойств «термодинамических единиц» – человеческих индивидов.
Отбор и приспособление сознательно временно исключены из предложенного описания, поскольку мы рассматриваем «борьбу за существование» как очевидный и радикальный фактор, в то время как нас интересует крайне слабый процесс, который хотя и может сопровождать примитивный отбор на неких начальных стадиях, но на самом деле «опережает» их и является исходным. Конечной целью инициативы является вызов «огня на себя», оживление полемики, привлечение различных знаний, дискуссии разных профессионалов.
Некоторые обстоятельства указывают на возможность количественного измерения тенденции к спонтанному росту сложности (полимеризации) в хаотических средах. Однако, не будучи профильными профессионалами, мы вынуждены ограничиться поиском подтверждений путем интерпретации известных фактов и обстоятельств. При этом мы хотели бы выслушать возражения со стороны различных научных школ и традиций. Возможно, лишь отдельные наши соображения найдут какое-либо подтверждение в дальнейшем, но и это было бы уже результатом.
4. «Step-фактор» или третий путь. Наша дополнительная идея связана с введением в описание базовых процессов молекулярной эволюции так называемого «шагового» (step) фактора. Решающее значение здесь имел не только процесс диспропорционирования энтропии, но и «внутренняя» тенденция роста сложности биомолекулярного материала, идущая малыми итерациям («N+1») и вызывающая обобщенный рост физического размера и уровня сложности базовых молекулярных агентов. Можно сказать, что это просто эвфемизм полимеризации в эмпирическом рассмотрении с учетом физико-химических основ этого процесса. Иными словами, элементарные модификации в исходном молекулярном материале имеют выделенное направление. Оно заключается в тенденции к накоплению размеров и сложности компонент. Тогда в рассмотрение включаются не только свойств специфических базовых процессов и реакций, конструкции искусственных организмов, но и обобщенные свойства возможных исходных сред и «целевых» выскомолекулярных смесей.
В экспериментальном сценарии «шаговый фактор» в прямом и преобразованном виде «управляет» всем дальнейшим ходом макроэволюции живого мира. Обобщение этой тенденции на крайне рассеяном и дисперсном материале и придание ему «физически» структурированного вида представляет значительную трудность, если такое обобщение вообще возможно. Мы попытались наметить для этого некоторые пути.
Данная конструкция может быть представлена в общем виде множителем Тинбергена egt, где g – темп прогресса какого-либо параметра живого (у Я.Тинбергена он символизирует технологическое развитие).
На реальность подобного подхода косвенно указывает эксперимент Миллера, создающий в мгновенном процессе аминокислоты и другие органических веществ (здесь следует назвать несколько имен – Д.И.Менделеева (возможность синтеза углеводородов в природных условиях), А.И.Опарина (высказавшего по существу идею эксперимента Миллера) и А.М.Бутлерова (синтез многообразия сахаров вплоть до рибозы В.Н.Пармона)). При этом мы просто записываем итоговое уравнение («получилось 22 аминокислоты»), в то время как этот неравновесный процесс прошел ряд этапов в скрытых переходных состояниях органического синтеза. Аналогичным путем могут быть получены и элементы кирпичей нуклеиновых кислот – азотистые основания. Получение нуклеотидов (на сегодняшний день получено только 2) проходит в цепи реакций, но требует исходной смеси компонент, что роднит его с миллеровским экспериментом. Подобная склонность к определенному типу структурных комбинаций может проявляться в различных формах и условиях.
Другими словами мы предположили, что фундаментальные процессы «не исчезают», а распространяются, нередко приобретая неузнаваемый вид. В соответствии с принципом своеобразного актуализма мы далее предполагаем, что макроэволюцией жизни управляют в фундаментальной основе те же силы, которые привели к ее возникновению.
5. Э.М.Галимов. Эта идея есть уже у Э.М.Галимова, но у нас она интерпретируется иначе – как тенденция к росту совокупной сложности (организованной и дезорганизованной компоненты), а не упорядоченности из беспорядка [1], чему внимание будет уделено несколько ниже (косвенно солидаризуется с идеей позиция [8]).
У Э.М.Галимова тенденция к упорядоченности, присущая процессам и явлениям неравновесной термодинамики (диссипативным структурам), неким образом проявляется в сложной организации, внутреннем соответствии цепей биологических процессов.
С нашей точки зрения эксперимент Миллера показал не только синтез определенной группы органических веществ, он показал возможность абстрактного процесса эволюции органических веществ в общем направлении биогенеза. Наблюдаемые более сложные конфигурации способны иметь свою скрытую «статистическую» причину, обусловленную на самом обобщенном уровне некоторой комбинацией химических сил. Таким образом, условная «внутренняя постоянная» – встроенная тенденция к росту сложности биополимеров – представляет собой эмпирическую констрyкцию, символизирующую потенциальную способность исходной среды эволюционировать в определенном общем направлении.
Длина генома человека составляет 2 метра, он состоит из порядка 3,5 млрд. пар нуклеотидов. Это означает, что он не мог не расти, начиная со странного состояния, близкого к «нулевому» размеру, усредненным темпом порядка одной нуклеотидной пары в год. В этой связи наша пробная логическая установка заключается в следующем:
1) Геномные молекулы и их исторические предки (если таковые существовали), а также их молекулярное окружение испытывают тенденцию к усложнению, выражающуюся в том числе в тенденции к росту их молекулярного размера;
2) Спонтанная тенденция к росту генома, если и не играет решающей роли в эволюции, то является автономным специфическим фактором роста сложности живых организмов;
3) Появившись на наиболее ранних этапах биогенеза медленная неустойчивая (статистическая), но «непреодолимая» тенденция к усложнению исходных биомолекулярных агентов в косвенном и опосредованном виде действуют в составе всех более поздних молекулярно-биологических процессов и систем.
Молекулярные единицы участвуют сразу в двух мирах, двух культурах – в молекулярной полимеризации и биогенетических гомеостазисах. Полионуклеатиды – это, с одной стороны, носители наследственной информации, а с другой – «обычные» высокомолекулярные полимеры, способные в определенных условиях расти дальше. Это обстоятельство позволило нам ввести понятие молекулярно-генетической системы, символизирующее «двойное дно» всей молекулярно-биологической реальности.
Наши предложения сводятся к приложению некоторого аппарата к изучению подобных «разделенных», «двусмысленных» систем. Среди двух традиций науки о происхождении жизни – метболизма и репликации [9] – можно наметить возможный третий путь, предполагающий самостоятельную роль структурного субстрата.
Обобщенная тенденция к усложнению обусловлена комбинационными свойствами элементов «углеродной группы» (С, Н, О, N, S, P и др.), вытекающими из их физико-химических характеристик. Эта очевидная идея периодически мелькает в научной и околонаучной среде. Мы попытались сформулировать ее несколько более рельефно и в акцентированном виде, позволяющем связать цепочку рассуждений с рядом фактов и наблюдений.
6. Свидетельствуют детали. По-видимому, принципиально не существует никакого способа оценить степень приспособительного качества того или иного организма (существование которого свидетельствует о его достаточности). Для того чтобы отделить причину и результат эволюции, необходим выход на уровень, где такое разделение возможно. «Внутренние причины развития» и даже наиболее радикальные ароморфозы, выглядящие как «превентивные эволюционные проекты», очевидно, могут быть лишь следствием неизвестных конкурентных сценариев адаптации, какими бы маловероятными они не казались. Ведь ни прямого, ни надежного косвенного доказательства здесь также не существует.
Однако дистанция между причиной и следствием может быть относительно различима в неких предельных случаях, например, на ранних стадиях существования и зарождении жизни, физико-химических и мысленных экспериментах, а также в определенных узловых эпизодах эволюционной истории. На роль подобной ситуации может претендовать «ископаемый» сценарий эукариотизации, состоявшийся в древнейшие геологические эпохи, особенности которого показал, в частности, известный российский биолог А.В.Марков [6]. Все дело в том, что рекомбинация органоидов в теле эукариота предполагала предварительное появление новых функциональных свойств геномного механизма. Без них «материнская» клетка не смогла бы «принять» органоидных симбионтов. Эта версия хорошо ложится на высказанную нами идею о превентивном и спонтанном характере роста генома, который обусловливает плавный переход от способности метаболических мутаций прокариотических организмов к уже эукариотической способности морфологических ароморфозов.
Это один из переходов сложности, который может быть отложен в системе когнитивных шкал (см. далее). Наша стратегия предполагает поиск в эволюционной истории узловых моментов, детали которых свидетельствуют в пользу того или иного подхода. На многих элементах сценария эволюции мы не можем остановиться в кратком материале, но они, как нам представляются, свидетельствуют о том, что постоянный внутренний источник роста сложности, «авансирующий» эволюционный процесс вполне мог существовать. Среди них – ароморфозы различных типов, которые могут быть отождествлены со скачками уровня организации биологических объектов, появление новых качеств и измерений степени сложности, компромиссный облик древнейших организмов.
Основой нашей экспериментальной стратегии является применение упомянутого выше принципа когнитивных шкал, когда возможные решения вычисляются методом экстраполяции в привязке к измерителям, выражающим предполагаемый смысл происходящих процессов. Комбинации «экспериментальных» когнитивных шкал и обобщенных свойств биологических объектов позволяют сформулировать новые подходы к сценариям и процессам жизни.
7. Абстракция сложности (T=0). Главная идея нашей комбинации заключается во введении внутренних измерений понятия сложности, в переходе от категории абстрактной упорядоченности (Э.М.Галимов) к категории абстрактной сложности как таковой. Сложность здесь – любая суперпозиция, взаимосвязанная комбинация более простых элементов. Ее абстрактная мера на базовом уровне физических процессов – «противотепловой» эффект стягивания, например, атомов в белковую глобулу. Подобная физическая мера (самая «простая» сложность) рассматривается как исходный пункт упорядочения. Любое хаотическое объединение элементов является «физически» упорядоченным, поскольку их положение не вполне произвольно. Простейшая сложность есть «минимальная» упорядоченность, однако сложность – не обязательно только регулярная упорядоченность и наоборот.
Сложность бывает организованная (упорядоченная) и дезорганизованная (хаотическая, включая «мусорную», «junk complexity»), каждая из которых играет свою роль в процессе взаимодействии этих абстрактных компонентов развития. Совокупная сложность достигает максимума при соотношении организованной и дезорганизованной («мусорной») компоненты в пропорции, например, пополам. Мы отводим этому феномену излишнего хаотического компонента геномов и других биологических (и не только биологических) структур особую роль.
Известно, что мусорная компонента генома человека достигает 97% его общего размера. Белки производят впечатление в целом хаотических объектов, содержательную роль в которых, на первый взгляд, играет лишь активная зона. Мы предполагаем, что «излишняя» хаотическая часть белка имеет по смыслу примерно тоже значение, что и мусорная компонента генома.
Избыточность – характерная черта такого явления, как жизнь. Она присутствует во многих микро- и макроскорпических биологических объектах и системах (см. п.7-8). Хотя «мусор» по смыслу не может быть изобретением дарвиновского отбора, он все же каким-то образом присущ процессам биологического роста.
Из предложенной цепочки рассуждений вытекают два сценария. Происхождение жизни могло протекать в двух вариантах. Речь идет о более привычном и традиционном «легком» и нашем парадоксальном «тяжелом» сценариях. Подобные соображения не являются новыми, но смысл традиционного легкого сценария заключается в том, что на ранних этапах действует своего рода маломолекулярный организм или его предок – объект, состоящий из малого числа малых молекул. Речь идет о неких минимальных бионтах, немедленно включившихся в естественный отбор; сами они также являются продуктом подобного отбора в рамках неких схем автокаталитических процессов. Таким образом, дарвиновская прогрессивная эволюция жизни может быть обращена вспять к самому началу t0, когда от эволюционной парадигмы остается лишь довольно смутная идея селективности. Легкий сценарий предполагает опытное получение базовых компонентов жизни и конструирование искусственных организмов.
«Тяжелый сценарий» заключается в ином – рубеж живого пройден с участием многомолекулярных и структурно сложных, довольно хаотических объектов и систем. Этот сценарий вытекает из идей спонтанного усложнения (step-процессы) и предполагает ряд важных деталей, на которых мы не сможем здесь останавливаться. Рост сложности стартовал ранее какого-либо аналога дарвиновского отбора в рамках сколь-нибудь развитых «правил эволюции», хотя мог включать селекционно-статистический фактор развития динамических систем, удаленными аналогами которого являются различные варианты известного селекс-процесса. Диаграмма, иллюстрирующая эту ситуацию приведена на рис.1; пунктиром «отсечено» появление настоящего генома.
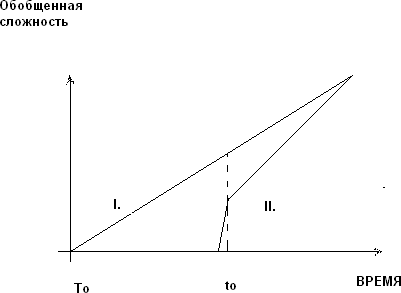
Рис.1. Схема «тяжелого сценария» эволюции.
В «легком сценарии» (II) живое после короткого неясного этапа добиологической эволюции очень быстро достигает степени сложности, достаточной для эволюционного отбора. Жизнеспособные объекты относительно просты, маломолекулярны и высокоупорядоченны, поскольку для «мусорной» компоненты в них просто не найдется места.
Можно сказать, что они уже «запрограммированы» однозначно для выполнения своей жизненной роли. Число биологических процессов в простом организме по тем же причинам, очевидно, не может быть велико. По логике, для появления простых структур не нужно много эволюционного времени, чем и объясняется их положение на временной оси диаграммы. Таким образом, сложность маломолекулярных организмов была невелика и затем быстро восполнена путем ускоренного отбора более сложных и поэтому более эффективных форм (рис.1). С нашей точки зрения «легкий» сценарий явно сталкивается с трудностью малых степеней свободы, которые недостаточны для любого подобия живого. В нем трудно найти место для обязательной мусорной компоненты. Более интересным кажется другой рамочный сценарий, в некотором отношении близкий к схемам Э.М.Галимова (рис.1).
В «тяжелом» сценарии (I) рост сложности начинается по обобщенным физико-химическим причинам, протекает относительно постепенно. Рубеж жизни пересекают уже достаточно крупные хаотические структуры со значительной «мусорной» компонентой, которая придает им вид телесных механических (механохимических) объектов. Их функционал хаотичен, но достаточно разнообразен. «Производство сложности» (complexity production) стартовало ранее гипотетических высокоактивных гиперциклов, ранее каких-либо преорганизменных объектов и тем более ранее появления линнеевских видов. В общем смысле хаотическая компонента, структурный мусор в исходных средах играет значительную роль, а в дальнейшем его аналоги обусловливают феномен опережающего развития. Умозрительные парадигмы тяжелого сценария по его логике выглядели бы столь необычно, что мы пока не будем их приводить и коснемся их ниже по тексту [10]. Тяжелый сценарий предполагает исследование и конструирование свойств смесей и обобщенных сред, массового поведения жизненных элементов.
Таким образом, интерес представляет область, близкая к появлению и началу роста настоящего генома (tо). Но существует и не совпадет с первой еще одна выделенная точка – момент начала роста обобщенной сложности (Tо<<tо). Эта конструкция оказывает влияние на рамочные параметры всего сценария жизни.
8. Абстракция роста (t=0). Какими могут быть эмпирические правила вблизи «особых точек» биологического роста? В дискуссионных целях можно сформулировать для них следующие эмпирические правила или свойства – инкрементальность, непроизвольность, неинвариантеность, геометризм, селективность, кольцевание и избыточность. Суммирует их аксиома опережения, перспективности, «ориентации на будущее» системы жизни.
1. Инкрементальность. Процесс биологического развития инкрементален на начальных этапах и в базовом виде осуществляется путем малых приращений – присоединением небольших мономеров и элементарных реакций сшивки длинных мономеров (что в целом можно рассматривать как «step», N+1).
2. Непроизвольность. Относительно более сложные молекулярные комбинации элементов и ранние циклические функционалы по смыслу обладают своего рода структурной («молекулярной») памятью [11] и ограничивают спектр возможных присоединительных и редуктивных «мутаций», предопределяя тем самым общее направление дальнейшего роста, последующую форму молекулярного объекта (и, в конечном счете, всей биологической системы).
3. Неинвариантность. Статистика неинвариантности прямого и обратного направления в процессах органического синтеза предположительной предбиогенной эволюции и дальнейших эпизодов эволюционной истории требует дополнительного изучения. Однако можно предположить, что случайные «прибавления» и «вычитания» в асимметричном объекте не вполне произвольны и ведут к эволюции и росту его структуры (в молекулах действуют выделенные области – «процепторы»; на смысл этого условного термина могут указывать особые свойства концевых участков ДНК и активные области других биополимеров). Эволюционный демонтаж генома в некоторых эпизодах видовой адаптации вел к сокращению архива признаков, но, по-видимому, никогда не вел к эволюционной редукции уровня и «стандарта» самого геномного механизма.
4. Геометризм. «Геометрия имеет значение». Жизнь это не просто цепь химических реакций, определяемая химическими свойствами биомолекул. С самого начала жизнь – телесная форма, механохимический феномен, где пространственные свойства играют существенную роль в накоплении сложности и функционального потенциала агентов жизни (механохимия – это, по- видимому, такая же «метка» жизни, как генный код). Чтобы обрести достаточное телесное измерение начальные агенты жизни должны были иметь «физическое измерение», т.е. достаточный размер и сложность. Уже самый начальный субстрат должен обладать механохимическими свойствами. Спонтанный рост и формирование хаотической компоненты – наилучший кандидат на причину телесного «наполнения» облика химических предков жизни.
5. Селективность. Более сложные объекты в составе организованной и хаотической компонент по статистическим причинам являются одновременно более реакционно способными и вариативными в обобщенном статистическом смысле. Тогда в случае включения в цепи циклических процессов отбор может «подкреплять» появление относительно более сложных структур, делая всю картину более контрастной. Проявлением этой тенденции с нашей точки зрения является известный селекс-процесс, в котром РНК in vitro самопроизвольно реплицируются и способны расти. С нашей точки зрения здесь происходит не что иное, как весьма энергичная генерация биоподобного материала путем полимеризации, репликации, а также «сшивки» концов РНК и т.д. (причем из-за невысокой точности копирования, скорее, «мусорной» компоненты) [12]. Из этого можно предположить, что аналогичные тенденции – высокая склонность к полимеризации и воспроизводству – возможны в других или аналогичных обстоятельствах и средах. Они могут сочетаться с процессами диспропорционирования энтропии, обратными связями по устойчивости «найденных» комбинаций, эволюцией гиперциклов. Во всяком случае именно в «селексе» нашло подтверждение наше понятие «шаг» и нет никаких оснований не придавать ему более широкого смысла.
6. Кольцевание. В условиях, когда спонтанный рост биологических структур идет по различным направлениям и в рамках различных групп процессов, их взаимодействие создает предпосылки появления и запуска сложных структур и функций. Кольца, то есть «неупрощаемые» функциональности и структурные конфигурации, как бы вырастают из спонтанно прогрессирующей среды. Иначе появление подобных образований, например, динамических «качений» компонентов рибосом объяснить, по-видимому, невозможно. Естественная идея селективности здесь играет свою роль, но не ранее чем для отбора созревают достаточные предпосылки.
7. Избыточность. Все организованные объекты избыточны, включают в себя дезорганизованную неупорядоченную компоненту, которая и является исходным «материалом» стохастического роста. Как известно, мусорная компонента человеческого генома составляет до 97% его размера. Вообще, сценарии раннего биогенеза и формирования химических предков жизни могут избежать избыточности, хаотичности и «ненужности» действующих агентов и элементов их структур. Поскольку хаотическая среда неизбежно будет мешать крайне тонкому маломолекулярному процессу, она должна быть в него каким-то образом включена. Cпонтанный рост по своему смыслу в высокой степени хаотичен, то есть избыточен, «излишен».
8. Рационированность. Любые чрезмерно энергетичные события способны нарушить систему слабых процессов и структур, поэтому метаболические параметры жизни нарастают инкрементально, что сопровождается равномерным распределениям энергетических перепадов по реакционным сетям (что отчасти и объясняет необъяснимую картину исключительно длинных и алогичных реакционных цепей). Высокая сложность несовместима с энергетическими градиентами, что не может не быть не связано с общими условиями исходных сред (тяжелый сценарий). Далее жизнь лишь постепенно выделяется из окружающей среды, и ее обобщенный метаболизм становится много энергичнее окружения.
9. Перспективность. Комплекс процессов и обстоятельств 1-8 порождает феномен превентивного развития, начиная с ранних и микроскопических фаз жизни и вплоть до согласованных комплексных эволюционных кампаний и ароморфозов высших организмов (так называемые «эволюционные проекты»). Из этого вытекает иллюзия или реальность «эвристических», проективных свойств генома. Биологические формы в эволюционном смысле имеют тенденцию как бы опережать окружающую среду, выходить на новые горизонты. При этом структура обгоняет функцию, в частности, когда геном растет «не зная, зачем ему это нужно», что отчетливо наблюдается в ряде основных процессов транспонирования генов, когда перенесенный ген приобретает новый функциональный смысл. Превентивной компонентой развития является мусорная (junk) часть генома и ее варианты. Аналогично генам избыточная и превентивная сложность химических предков жизни как бы постфактум «осваивается» ее функционалом.
В комбинации с правилами идут основные эвристические шкалы:
- Шкала сложности и ее компонентов;
- Шкала степени локализации и обособления организмов (включая геномную, половую, иммунную, морфологическую и др.);
- Шкала силы катализа и уровня метаболизма, меры термодинамического порога;
- Шкалы частоты мутирования, смены поколений, размеров и возраста организмов.
На пересечении подобных шкал отмечается появление новых биологических объектов и парадигм, эпизоды сценариев эволюции. Совокупное развитие не может не носить колебательного или периодического характера, порождая границы этапов общей организации жизни. В частности, предел сложности означает ситуацию, когда дальше сложность может расти только в другом, новом измерении, так как это было в случае эукариот (переход от сложного метаболизма к сложной морфологии) и ряде других аналогичных ситуаций. Не расти сложность не может, поскольку это задано свойствами биомолекулярных систем и живого вещества в целом. Вперед общий сценарий продвигается благодаря нашему гипотетическому step-процессу.
9. Латентный катализ. Весьма ярким свидетельством возможностей предложенной логики является полученный молекулярными биологами процесс «холодного синтеза» полинуклеотидных цепей из негеномных нуклеотидов без минеральных подложек, автокатализа и «диспропорционирования энтропии» [12]. Дальнейший путь решения задачи, очевидно, может состоять в постепенной подстановке негеномных нуклеотидов геномными в рамках какого-либо поэтапного процесса. При этом «нулевой» катализ может полагать начало структурной шкале силы катализа вблизи особых точек T0 и t0, вблизи исходного термодинамического равновесия (Smах).
Читаем в литературе: «Многие ученые начинают склоняться к идее, что вместо относительно сложных азотистых оснований, присущих современным нуклеиновым кислотам, их предшественницы могли использовать слегка измененные варианты молекул, более склонные к реакциям нематричного синтеза» [12]. Это прямое указание на возможное существование целого мира процессов при слабом катализе и иных условиях, способствующих спонтанному росту предбиологического субстрата. Ведь практически никто не мог предвидеть колоссального класса молекулярно-биологических процессов, до того как они были наблюдаемы экспериментально. В молекулярной биологии существует указание на такую возможность в виде гибридных, переходных и модифицированных форм. Впрочем, мир подобных объектов должен значительно превосходить ныне известный.
С физико-химической точки зрения эксперимент Миллера – это группа довольно специфических химических реакций. С эмпирической точки зрения это тенденция, способность определенной группы веществ к трансформации в предбиологическом направлении, встроенная склонность к образованию химических предков жизни. С абстрактных позиций это есть скачок сложности, структурной организации.
В этой связи в абстрактном виде существуют две формальные возможности:
1) Вторичный самовозникающий процесс роста существует «физико-химически». При этом он в чем-то аналогичен, например, цепным реакциям, обусловленным микроскопическими свойствами реагентов и макроскопическими условиями. Однако он обладает некоторыми специфическими особенностями – крайне низким темпом, жесткой обусловленностью внешними условиями, вероятностной природой и пр.
2) Если вторичные процессы и явления как таковые не существуют, то обобщенные с помощью того или иного приема физико-химические характеристики молекулярной плазмы, ведущие к ее усложнению под действием факторов не физико-химической природы (например, отбора, гиперциклов и т.д.), могут быть различными путями включены с системное рассмотрение.
Компромисс этих подходов довольно ярко высвечивает проблему трудностей научной логики на пересечении нормативных и эмпирических оснований естественных и социальных наук. В общем смысле асимметрические свойства высокомолекулярных смесей отрицать, кажется, невозможно. В том или ином виде step-process (как, например, «мутации роста») не может не существовать; вопрос только в степени его обусловленности и роли. Очевидно также, что в «атмосфере» катализа тенденция к полимеризация возрастает. Здесь просто введена некая новая шкала, остальное покажет апробация.
Если программа редуцирования жизни до элементарных зародышевых комбинаций и впредь не окажется продуктивной, то не останется никакой иной возможности, кроме перехода к стратегии холизма – изучения не свойств изолированных объектов, а обобщенных процессов и их производных. Из нашего подхода следует, что современный облик биосферы в целом не случаен, а скорее, предписан параметрами базовых компонент, которые по статистическим причинам могли реализоваться только в некотором определенном виде.
Мы стремились дать свое краткое определение жизни. Сильно утрируя, можно сказать, что исходные элементы жизни как бы «хотят объединяться», но этому препятствует обобщенный потенциальные барьер. Обобщенный катализ его ослабляет и образуется единое историческое сооружение –жизнь. Производство этого катализа требует средств, поэтому жизнь – это еще и тепловая машина, для работы которой необходима энергия различных видов.
10. Последний аргумент. В эволюции живого все немного растет: известны эффекты обобщенного роста размера генома, числа хромосом, размера белков, их разнообразия и количества, разнообразия тканей и клеток, количества клеток. Происходит рост размера и возраста организмов, числа нейронов, «потока» синапсов, их организации и разнообразия. За этим следует экономический рост, социально-политическое развитие и технологический прогресс.
Э.М.Галимов интерпретировал «формулу жизни» как воспроизводство исходных компонентов путем действия созданных ими самими стандартных промежуточных агентов («большой гиперцикл»). Мы предлагаем ввести внутреннюю постоянную или инкрементальный фактор роста, проявляющийся уже в фазе исходной минимальной модели. При этом формула компонентов жизни претерпевает постоянное спонтанное усложнение («N+1»).
По нашей мысли существует тенденция не просто случайного мутирования генных кодов, но и случайного его приращения на каждом следующем этапе. Стандарт матрицы, медиаторов, общего облика структур и функций имеет тенденцию «все время немного расти» во всех пунктах и элементах биологической системы.
Формулу Галимова, выражающую самовоспроизводство компонентов жизни, можно дополнить условной поправкой [1]:
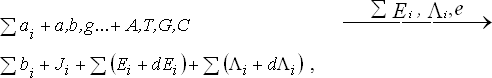
На ранних этапах малые приращения выражают образование «мусорной» компоненты (junk-factor), символизирующего одновременно эволюцию физического облика жизни и начало морфологии живого. Априорная сложность, в конечном счете, имеет эволюционный смысл и становится фактором естественного отбора.
В этом заключается смысл идеи «внутреннего притяжения» или «эмпирической постоянной» (step-процесс). Ведь элементы молекулярно-генетической системы при всех обстоятельствах остаются физико-химическими объектами, испытывающими воздействие рассеянной дроби («газа») элементарных модификаций (можно сказать – «клеточное тепло»). В развитом организме они «управляемы», а неуправляемые – в основном фатальны. На ранних стадиях, для которых главным образом и сделаны эти допущения, данный обобщенный статистический процесс был (предположительно) основным «спонсором» спонтанного роста.
Особенность нашего подхода заключается в том, что главной абстракцией биологического роста признается квазилинейная инкрементальная тенденция, создающая «частные случаи» экспоненциального, гиперболического (включающего «обострения») и спирального развития.
Математикам известна L-модель, которая с нашей точки зрения является перспективным кандидатом на метафору предложенного процесса «роста-компактификации», «свертки» развития, биологической эволюции в целом. Речь идет о «двойной спирали» размера и уровня сложности, роста «хаотического размера» и структурной организации. Дело в том, что L-функция создает два пространства – расширения и сжатия, представляя собой компромисс степенной функции и экспоненты [13]: ns=esln(n)
Аналогичные интерпретации содержатся в концепции системогенетики крупного российского специалиста в области квалиметрии и системных исследований А.И.Субетто. Здесь мы дополняем эту доктрину, в частности, фактором «мусорной» компоненты, шкалами организации и дезорганизации совокупного роста, а главное, пытаемся усмотреть специфическую «механическую» причину и некий глобальный термодинамический контекст.
Рост есть функция общности, тенденции к ассоциированию термодинамических единиц объединенного процесса... Мы создали экспериментальную конструкцию главным образом для анализа деталей научной логики, оказания воздействия на качество «стандартной модели», в которой наблюдается немало логических пробелов.
Тогда в определение жизни вносится новый мотив. Это есть комплекс процессов, обусловленных с тенденцией группы химических веществ к спонтанному усложнению, т.е. их фундаментальными физико-химическими свойствами. Это своего рода семантическая («речевая») структура (если можно так выразиться, «разговор природы»), стилизованная в рамках своих обобщенных статистических свойств.
Геномные паралогии (смены функционального смысла без изменения формулы) для нас говорят о многом. Если такое переформулирование возможно в молекулярно-генетической системе, то оно в каком-либо виде возможно и в догеномной материи. Кстати, оно не имеет отношения к химии, где изменение формулы не может не изменить функционал!
Наш последний аргумент – это кварки. Для них не существует принципа суперпозиции, вернее существует принципиальный барьер для кварковых комбинаций, за исключением нескольких, разрешенных законами великого объединения. Предел сложности существует и для сложности атомов (радиоактивный распад), молекул (термодинамический фактор).
Возможно, кварки – это начало глобальной шкалы сложности. Далее вселенная «диспропорционирует» ядра и атомы легких, затем тяжелых элементов, большие и малые органические молекулы..., возможно, короткие и длинные полимеры.
C точки зрения физики взрыв сверхновых порождает тяжелые элементы и энтропию, а с точки зрения смысла – фактор суперпозиции, новую сложность. Если «процесс сложности» как парадигма суперпозиции носит фундаментальных характер, то в нем запрограммирована жизнь. Хотя физик В.А.Эткин считает, что неравновесные процессы во Вселенной взаимоуравновешивают друг друга, нам представляется, что это единая цепочка. Не заметить ее нельзя.
- Галимов Э.М. Феномен жизни. М.: 2009.
- МакКонки Э. Геном человека. М.: 2008.
- Еськов К.Ю. Удивительная палеонтология. М.: 2007.
- Мейен С.В. Следы трав индейских. М.: 1981.
- Гринин Л.Е., Марков А.В., Коротаев А.В. Макроэволюция в живой природе и обществе». М.: 2009.
- Марков А.В., Коротаев А.В. Гиперболический рост в живой природе и обществе. М.: 2009.
- Материалы сайта (www.macroevolution.narod.ru).
- Эткин В.А. К термодинамике биологических систем (http://zhurnal.lib.ru/e/etkin_w_a/kenergodynamikebiologicheskihsistem.shtml).
- Шапиро Р. У истоков жизни (http://www.sciam.ru/article/3474/) 15.10.2010.
- Богоцкий С.В. От молекулярного ламаркизма к дарвинизму// «Природа», №11, 1990.
- Пармон В.Н. Новое в теории появления жизни (http://vsyachin.ru/earth_sciences/life_genesis_3.html).
- Григорович С. Вначале была РНК? В поисках молекулы первожизни// «Наука и жизнь», №2, 2004.
- Абраров Д.Л. Зеркальная симметрия и кольца инвариантов. Как строится математическая политика государства (http://www.chronos.msu.ru/RREPORTS/abrarov_zerkalnaya.pdf).


























Написать комментарий
Вы прямо не вбровь, а в глаз, уважаемый г-н Оксанов.Все так и есть. Есвенно это необходимо разглядеть. Для этого необходима немного необычная, хотя и очевидная логика. Я уже об этом напсал.Думаю, где и как опубликовать. А пока события развиваются быстро. Открыты дополнителые функции лишнего генома. Это и следовало ожидать. Это есть в статье. Сложные инрактивные циклы. Геном это почти компьютер."Первые мозги". Обосновано предположить, что проекты почти вычисляются. Предпроектируются. Против чего сильно возражает в преписке и в публикациях А.В. Марков. Эволюция нас рассудит.
Ау, чего коменты не доходят?
В экономике действует принцип расширенного воспроизводства ресурсов. При этом ресуры не только расширенно воспроизводится количественно, оставаясь неизменными качественно, но расширенно воспроизводятся качественно, когда исчерпание одной разновидности ресурса приводит к появлению новой разновидности, обычно, более сложной. Это хорошо видно на примере развития сырьевых ресурсов, ресурсов средств производства (усложнение интрументов, применяемых в экономике), и не только (от дров к атомной энергетике). Своего рода процессы, направленные против увеличения энтропии, естественно, только на ограниченном участке мира. Мне представляется, что в этом есть нечто, подобное биологическому развитию. Вероятно, такие процессы происходят и в сфере социального развития. Природа едина? Тогда и в экономике, и в социологии должно быть и много "мусора". Ваше мнение? Оксанов
По прошествии некоторого времени могу сделать только один комментарий. Ричард Докинз считает, что дать определение понятию "индивидуальный организм" невозможно. Не сущесвует. см. Ричард Докинз. "Расширенный фенотип". М.2011. Конкурируют не столько организмы, популяции и виды, сколько "расширенный фенотип". Это меняет многое и полностью вписывается в приведенную логику. Прогресс жизни тогда -- не столько обособление организмов, сколько структуриррование живого, обособление как таковое, стпень корого возрастает.
Уважаемые читатели! Как выясняется чрезвычайно близкие мысли были высказаны F.Spier в F.Spier Structure of Big History(1996). Резюме в виде статьи недавно опуликовано в одном из сборников под редакц.в т.ч.А.В.Маркова "Большая история: энергия, энтропия и эволюция сложности"."Эволюция, аспекты современного эволюционизма" M.URSS.2012